Mendelian Genetics
To Pea or Not to Pea: Mendelian Genetics
Gregor Mendel (1822–1884), an Austro-Hungarian monk, discovered what is considered to be the very foundation of genetics by growing pea plants in the garden of his monastery. He didn't have lab or sophisticated equipment, no exotic islands with unusual species to inspire him, just some dirt, some pea plants, and of course, lots of curiosity. However, he never experienced his own fame: his contributions were completely ignored or forgotten until the 20th century.Peas are wonderful little things. They are loaded with vitamins (as you have probably heard many times), but many other characteristics made them ideal for Mendel's experiments on the laws of heredity. Pea plants can be grown in large numbers, have a short generation time, and individual plants have both male and female reproductive organs so that they can self-pollinate or reproduce as either male or female. Finally, it is possible to control which individuals mate by simply transferring the pollen from one plant to another with a brush (cross-pollination).
Recap: Plant Reproduction
The male reproductive part of a flower is known as the stamen, which consists of a pair on anthers attached to a stalk known as the filament. Cells within the anthers divide by meiosis to produce the male gametes (sperm), which end up stored inside the plant's pollen.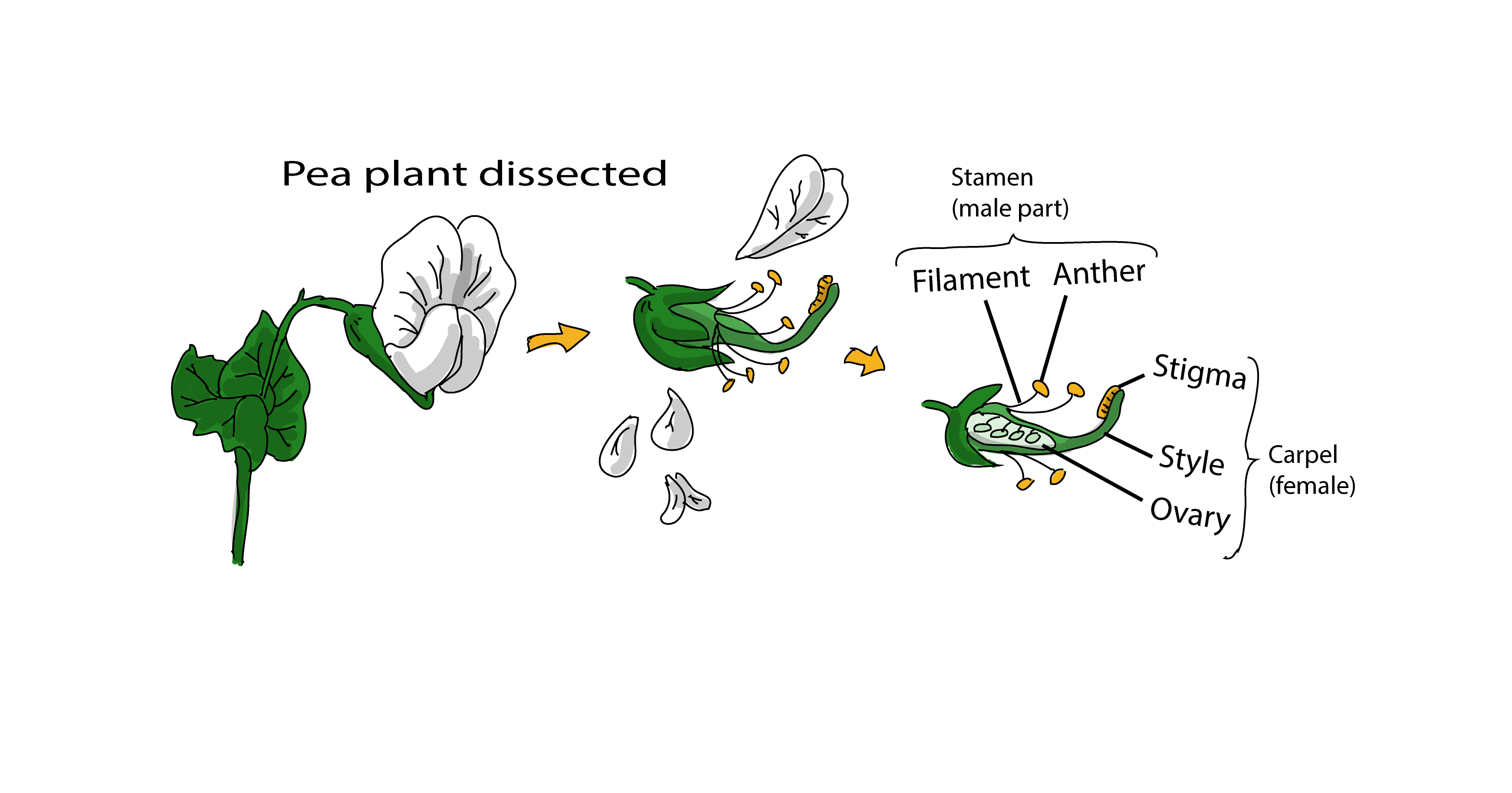
The female reproductive part of a flower is known as the carpel, which consists of a stigma, an ovary and style (which connects the stigma and ovary together). Inside the ovary there is at least one ovule, where the female gametes (eggs) are formed. The stigma catches pollen from the stamen, which then germinates and grows a tube down through the style to the ovules, enabling the plant's sperm to be delivered to a waiting egg, where fertilization takes place. In pea plants, these fertilized embryos go on to become the seeds (peas) contained inside a protective pea pod that formed from the ovary.
Geno and Pheno
Before we go any further into Mendel's fascinating discoveries, let's cover some essential terms. The characteristics of an individual that are inherited by children from their parents are called hereditary traits. Traits are genetically determined: segments of DNA called genes control them.The genetic constitution of an organism is called the genotype, while the phenotype is the observable manifestation of the genotype. So in the pea plants, a phenotypic characteristic might be flower color or height, while the collection of genes determining these characteristics is the genotype.
Mendel studied seven different phenotypic characters in pea plants, each with two forms or traits, including seed shape (round or wrinkled), seed color (yellow or green), flower color (purple or white), and stem height (tall or dwarf). Using a brush, he manually transferred pollen from one parental plant to another. These plants were called the parental generation, or P.
In the parental generation he had only true-breeding plants (true-breeding plants present exactly the same trait throughout many generations). No matter what plants he chose to breed, every single time the offspring looked exactly like their parents. These offspring, or "children," are known as the first-filial generation, or F1. Mendel recorded the phenotype of each F1 plant (loads and loads of them) before allowing them to self-pollinate to generate the "grandchildren": the second-filial generation, or F2.
The Monohybrid Cross
The most basic experiment, a monohybrid cross, involves two parental plants true-breeding for a single character but each carrying a different trait: for instance, two plants bearing different seed shapes (a character), one round and the other one wrinkled (two different traits). If "selfed," or allowed to produce seeds using its own eggs and sperm, each plant will produce either round or wrinkled seeds because they are true-breeding. But how about when crossed with one another? To figure this one out, we need to examine what kind of gametes (eggs and sperm) each of our plants produces.Most plants, like humans and most other animals, are diploid: they have two copies of each chromosome and thus two alleles, or copies, of each gene (even if that particular gene can have two, three, four or even more different alleles). Alleles are different forms of the same gene: in this example, one allele for seed shape produces round seeds; the other allele produces wrinkled seeds. In each gamete (egg or sperm), there is only one copy of each gene or, to put it another way, a single allele. Gametes are thus considered haploid.
Two gametes fuse together to produce a new diploid individual. How do gametes get to be haploid in the first place? Diploid cells undergo meiosis and in the process produce haploid gametes. What allele goes into each egg is determined by chance: 50% will have one allele, 50% the other. In a way it is like a bag of marbles holding equal numbers of two kinds of marbles. For each gamete, one marble is drawn at random from the bag.
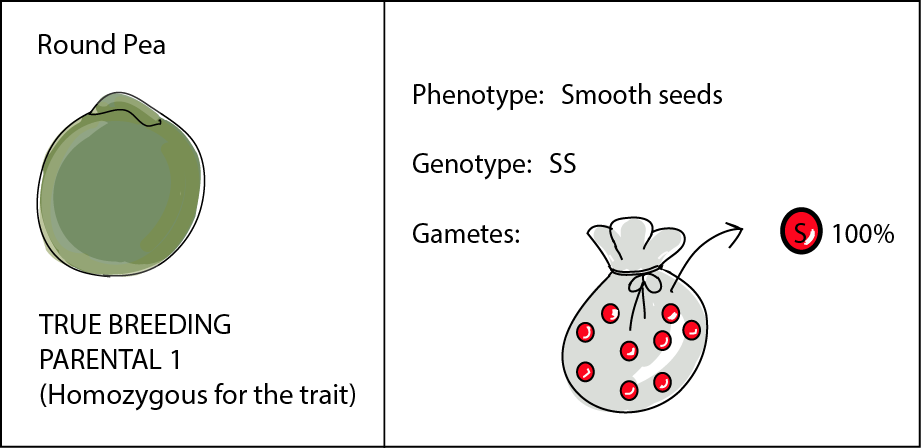
Our pea plant that produces only round, smooth seeds has a genotype (or genetic constitution dictating seed shape) that we denote as SS. Note: it really doesn't matter what letter you give to your alleles, just so long as you make a note of what they represent. But you do need to make sure that you stick to the same letter for both—in this case, S for smooth and s for wrinkled.
So at the locus for seed shape (the genetic "address" or location of seed shape), this pea plant has two identical alleles for smooth seeds. Because this plant is true-breeding, both alleles are the same (S), so this plant is considered homozygous. So this plant's bag of marbles contains only one kind of marble and can only produce gametes carrying allele S.
If this pea plant would get carded, this could be its ID:
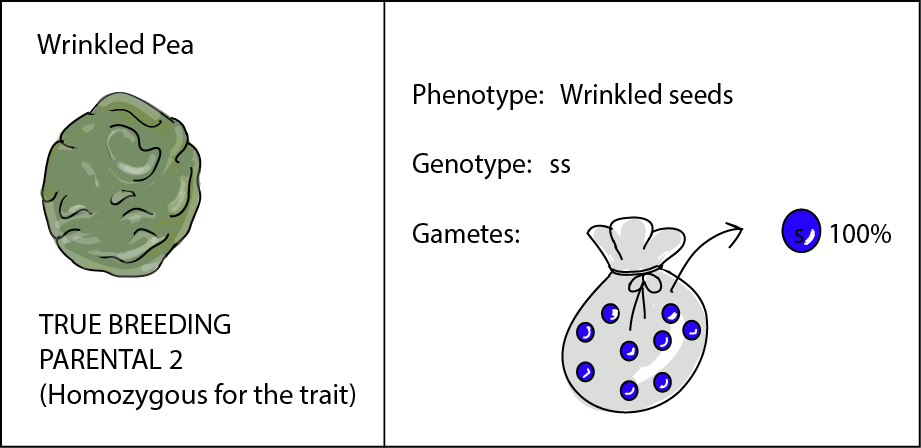
Our pea plant that produces only wrinkled seeds has a genotype of ss. Because this plant is true-breeding, both alleles are the same (s), and this plant is also considered homozygous. So this plant's bag of marbles contains only one kind of marble and can only produce gametes carrying allele s. Here's its ID:
We have the profile on our parental plants. Now, let's cross them:
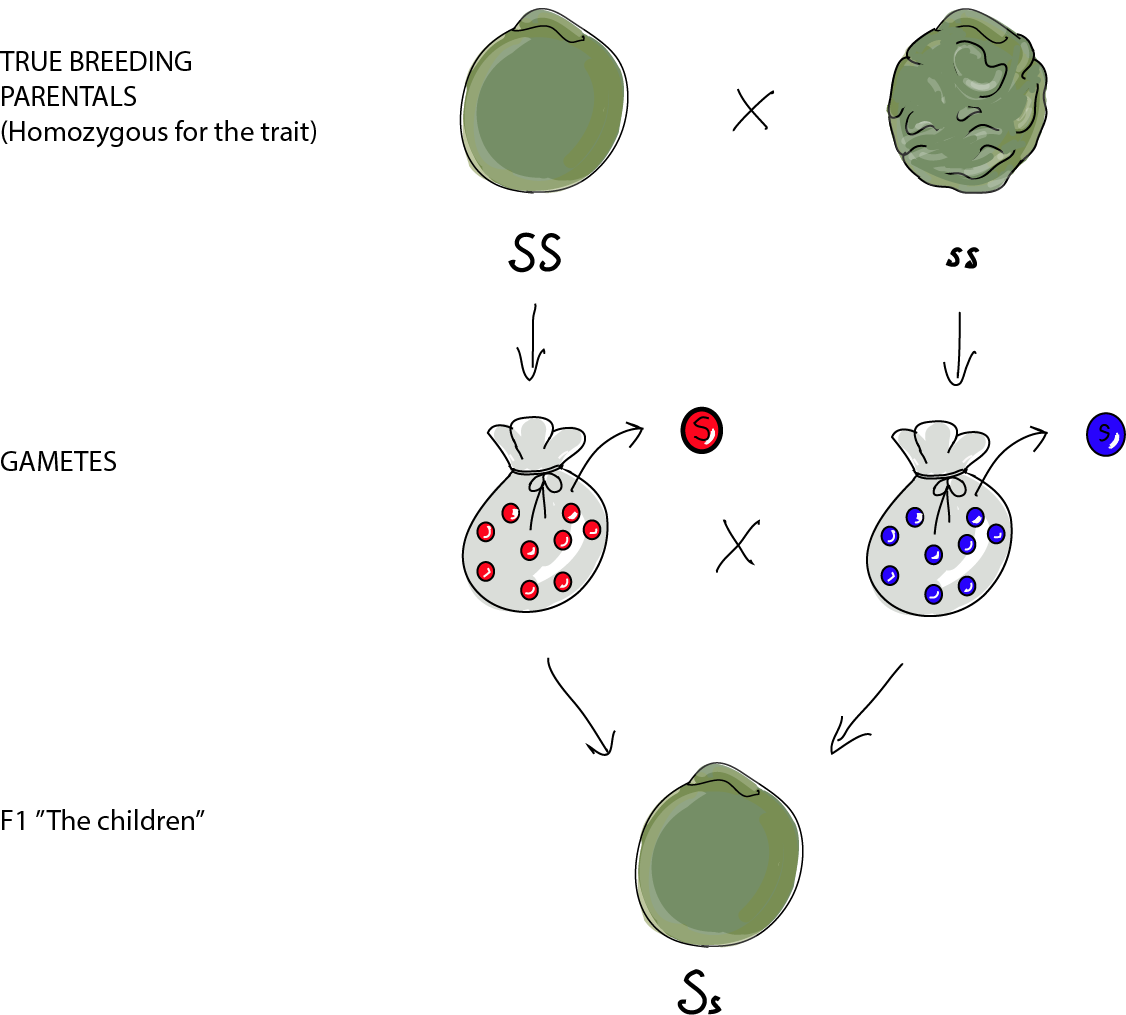
The F1 (the "children") all have the same genotype of Ss, because they drew an S marble from the round-seeded parent and an s marble from the wrinkled-seeded parent. All of the F1 plants produced round, smooth seeds like Parental 1. They are heterozygous for seed shape as they have two different alleles at this locus: an S allele and an s allele. When it comes to the F1 producing their own gametes, unlike their parents, they produce two different kinds of gametes, about half bearing an S allele and the other half an s allele.
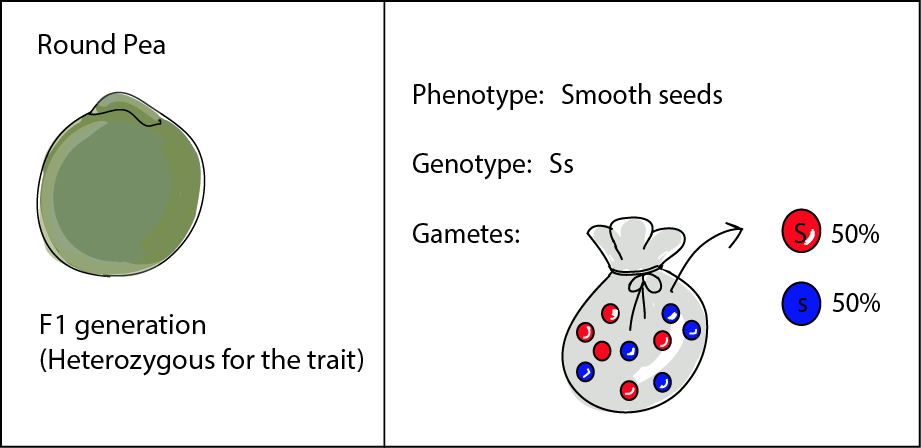
Mendel then took 253 of these F1 seeds, planted them, and allowed the resulting plants to self-pollinate to produce a new generation: the "grandchildren" or F2. The F2 were a different deal from their F1 parents. Out of the 7,324 seeds produced, 5,474 were smooth, while 1,850 were wrinkled. So there were a lot less wrinkled seeds than smooth ones. The ratio of 5,474:1,850 is, in fact, pretty much 3:1. There were three times as many smooth seeds as there were wrinkled ones. This is known as the phenotypic ratio because it looks at the relative numbers of each of the phenotypes.
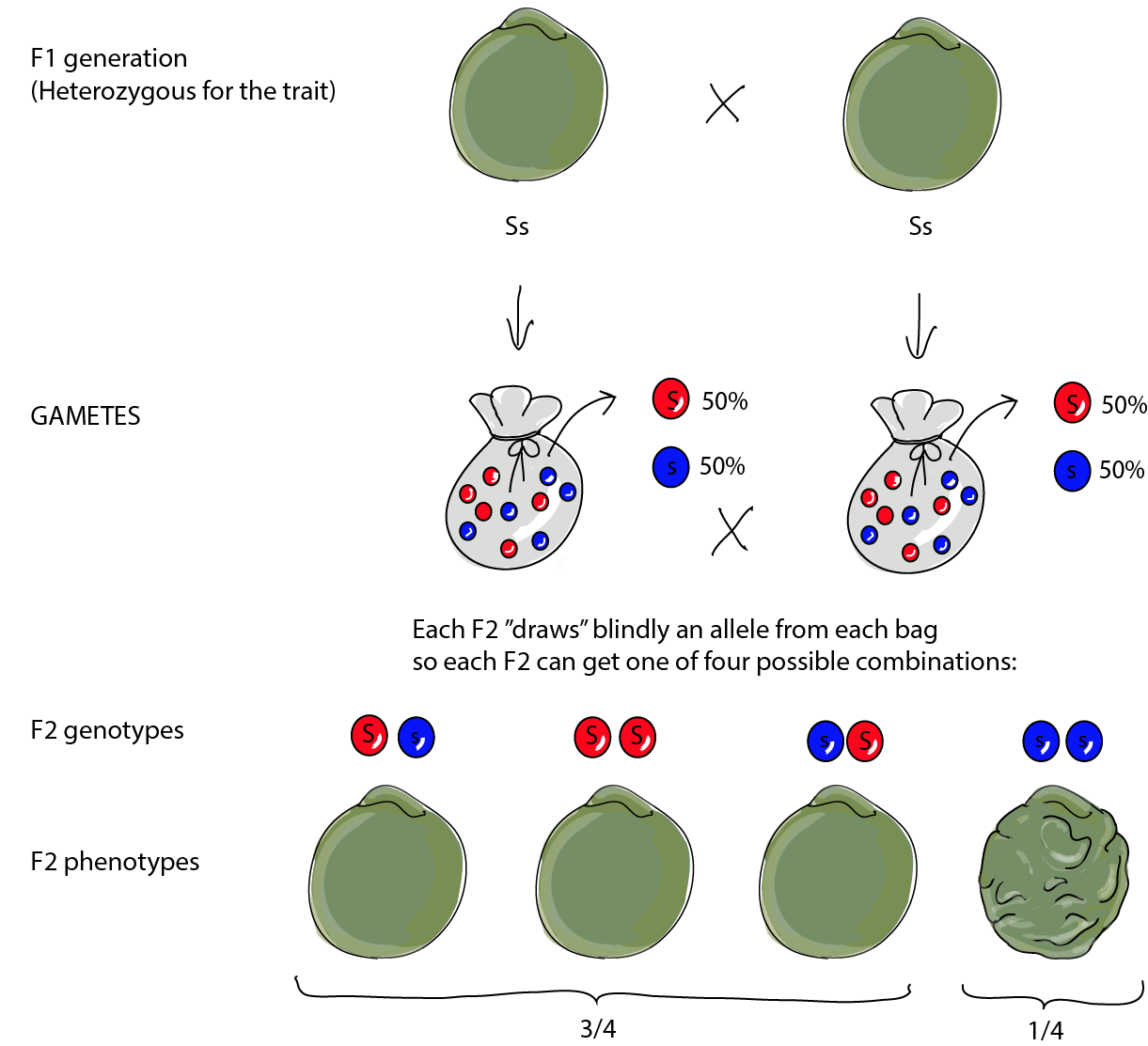
Don't forget that when you're writing out heterozygous genotypes the capital letter always comes first!
Let's review Mendel's observations. When crossing a true-breeding plant that produces round seeds to a true-breeding plant that produces wrinkled seeds, all the "children," or F1 plants, look like the round-seeded parent. When interbreeding the F1, ¾ of the "grandchildren," or F2, have smooth seeds, while only ¼ have wrinkled seeds.
Because both the true-breeding, homozygous plants (SS) and the heterozygous (Ss) plants show the round-seed phenotype, the round form of the pea is called the dominant phenotype, which is coded for by the dominant allele, S (dominant alleles are always represented by a capital letter). The wrinkled form is considered recessive. Only true-breeding, homozygous plants (ss) present this phenotype (and the recessive allele, which codes for that phenotype, is always represented by a lowercase letter).
The F1 generation are all heterozygotes (Ss) and thus always present the dominant trait. The recessive trait, wrinkled seeds, reappeared in F2, but only in a fourth of the plants because there are many more that are either homozygous dominant or heterozygous (and are, therefore, round seeded).
The Law of Segregation
Mendel studied seven different traits in pea plants, and in all those cases, he consistently observed the same results. From all these data, he derived the first law of Mendelian genetics: the law of segregation. When an individual produces gametes, the two alleles (the different forms of a gene) it carries go their separate ways or segregate, so each gamete receives only one copy.To put all this together, the true-breeding parents can each generate only one kind of allele, either smooth (S) or wrinkled (s). The F1 all have the same genotype, Ss, and since the S allele is dominant, these individuals all look the same (that is, round). The F1 generation, however, can produce gametes carrying both types of alleles, S and s.
If we go back once more to our bag of marbles parallel, the bag of marbles for an F1 contains both S marbles and s marbles in roughly equal numbers. Therefore, if you just dip your hand into that bag and draw a marble at random, about 50% of the time you'll get an S and 50% of the time an s. When these are combined again to produce the F2, all possible combinations of alleles can occur: SS, Ss, and ss.
Look back at the genotypic ratio for this cross. Did you notice that it's different from the phenotypic ratio? There's one SS, two Ss and one ss, giving a ratio of 1:2:1. This difference is caused because both SS and Ss, even though different genotypes, produce exactly the same phenotype. So ¾ of the seeds look round and only ¼ look wrinkled (our 3:1 phenotypic ratio, just written in a different form).
Punnett Squares
Punnett squares—named after another early geneticist, Reginald Punnett—are a great way to examine the genotypes and phenotypes resulting from a cross. They are the best way to keep your phenotype and genotype ratios neat and organized. The experiment we were just discussing can be analyzed with a simple diagram rather than a few paragraphs.On a simple square, we add one parent and its corresponding gametes to the length, the other parent and its allele to the height:

By simply filling each cell of the grid with the combination of alleles from top and side, we know all the possible genotypes in the F2:
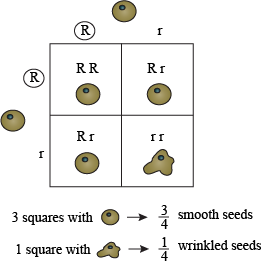
We can also quickly derive the ratios of each genotype and phenotype—it is simply a matter of counting. One out of our four cells contains the SS genotype (¼ SS), two out of four cells contains the Ss genotype (¼ + ¼ = ½ Ss), and one out of four cells contains the ss genotype (¼ ss). We're back to our 1:2:1 genotypic ratio! As you can clearly see, there are three round seeds to only one wrinkled one (our phenotypic ratio).
Mendel, as a good scientist, confirmed his conclusion about independent segregation using a different method with specific predictions: a test cross. In this case, he crossed a heterozygous plant with a homozygous recessive plant. He predicted the next generation would show a 1:1 ratio of round to wrinkled:

In this case, we have two out of four cells that are Ss (¼ + ¼ = ½ Ss), resulting in half of the plants producing round seeds. Also, we have two out of four cells that are ss (¼ + ¼ = ½ ss). As his predictions held true, he was able to support his original conclusions.
The Dihybrid Cross
We can now draw a Punnett square to show the possible genotypes and phenotypes for a single characteristic. Mendel didn’t stop there, though. Pea plants have lots of traits, and he also investigated how multiple traits were passed from parent to offspring. This is the infamous dihybrid cross.In a dihybrid cross, the parental strains are different for two characters: for example, seed shape (round, wrinkled) and also seed color (yellow, green).


As you may expect, now that we're dealing with two separate characters, things get a bit more complicated. The F1 all look the same again, round and yellow, and they're heterozygous for both traits.
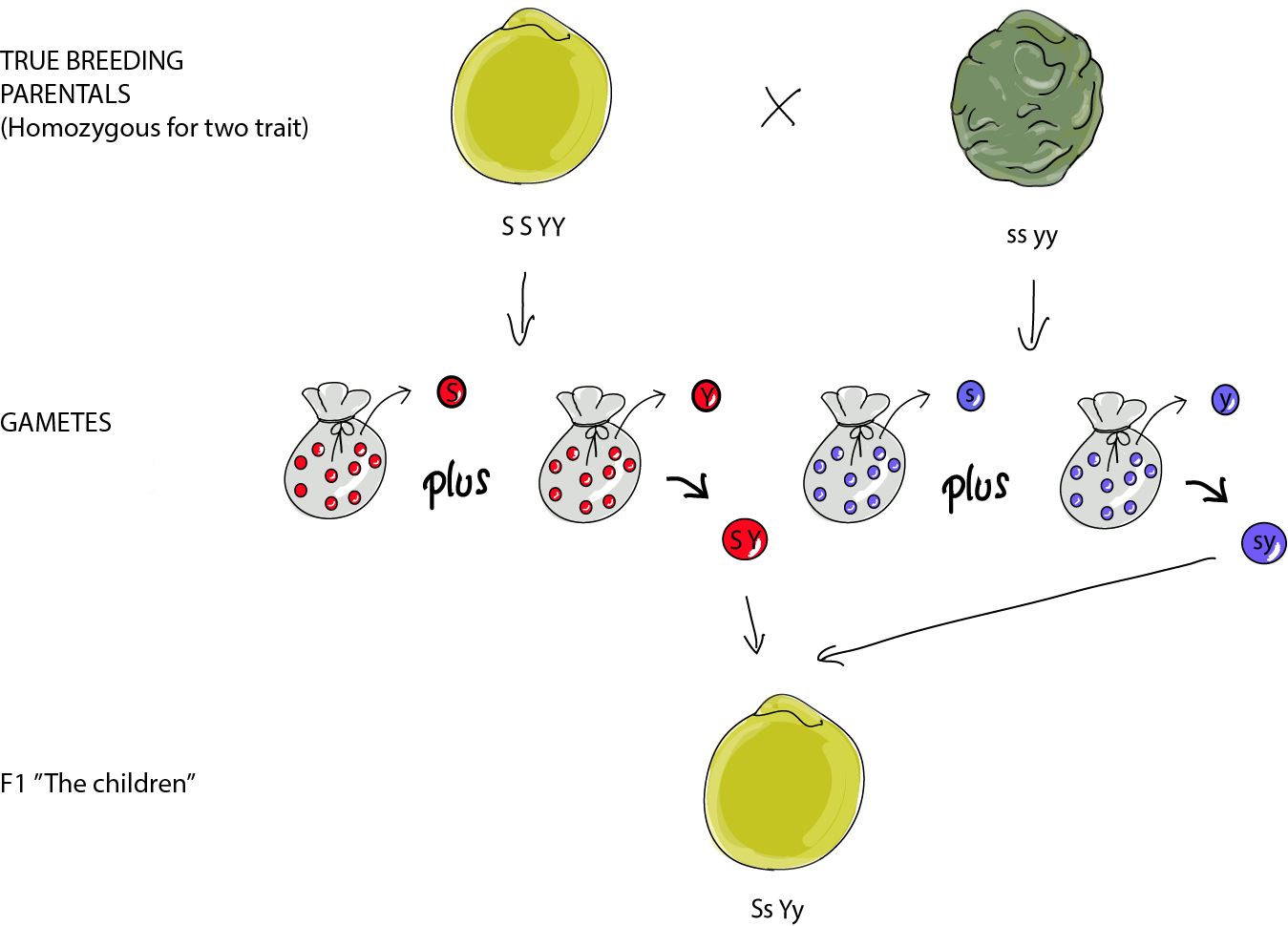
Here is the ID card for this dihybrid F1. For the gametes, think again of bags of marbles. Each locus (again, the genetic "address" or location for the character in question) has its own bag, so each gamete takes a marble from each bag. In the seed shape bag, we have S and s marbles in equal numbers. In the seed color bag, we have Y and y marbles in equal numbers. For each gamete, fate just dips its hand in each bag to pick a marble for each gamete.

Now, the F1 are selfed to produce the next generation, the F2. It gets kind of complicated to think through a diagram with arrows and bags when you have more than one locus, so a Punnett square is definitely the way to go!
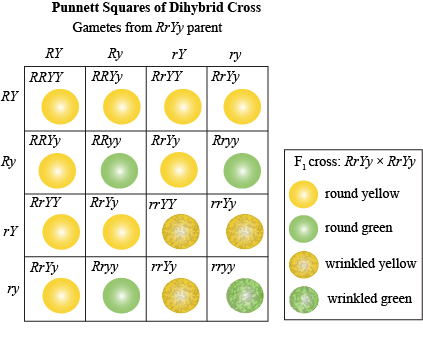
The F2 can have four different phenotypes combining the four different traits: round and yellow, round and green, wrinkled and yellow, wrinkled and green. And after counting hundreds of F2 seeds, Mendel noticed that once again he had a predictable, repeatable ratio of the different combinations. This time it was 9:3:3:1.
The Law of Independent Assortment
The dihybrid cross led to the formulation of the second law of Mendelian genetics: the law of independent assortment. During meiosis or gamete formation, the segregation of alleles for one character is independent of the segregation of alleles for another character.It's really important to remember that this is because the characters Mendel studied (and that we're discussing in this section) are located on separate, different chromosomes called non-homologous chromosomes. During meiosis, each chromosome from a homologous pair makes its way into a gamete.
Remember, in a diploid organism there are two sets of chromosomes, so each chromosome has a homologue—that is, a copy of itself, one of which you inherit from mom and the other from dad. Exactly which chromosomes from the different homologous pairs are sorted together into the same gamete is determined by random chance (aka luck), so characters on different chromosomes are inherited independently of each other.
In our marble bag story, all of this boils down to each gene/character having its own bag. A gamete needs a marble from each bag. Because the marble drawn from the first bag has no effect whatsoever on what marble is drawn from the second bag, whatever marble comes out of each bag is an unrelated, independent event.
Brain Snacks: Examples of Mendelian Genetics in Humans
Looking for a way to spice up the conversation when your whole extended family comes over for Thanksgiving dinner? Impress them with your snazzy new genetics knowledge!
- Wet or dry earwax: Yep! Next time you scratch the inside of your ear, you should look at the consistency of your earwax with your "scientific lenses" on. And, if you dare, ask some family members about theirs. In this way, you could do a study on a human Mendelian trait. If you manage to get over the "yuck" factor, you may notice that this somewhat icky trait can generally be classified as "wet" or "dry," and the "wet" phenotype is dominant.
- Attached or detached earlobes: Okay, okay, so maybe earwax consistency is a bit too much for you. How about another ear characteristic instead? Whether earlobes are "attached" or "detached" is another trait that some people believe is inherited like a Mendelian trait, where "detached lobes" is the dominant phenotype.
- Albinism: The amount of melanin, an amino acid-based pigment, helps to determine skin, hair, and eye color in humans. People that present with albinism do not have any pigment because they do not produce melanin. Albinism is always recessive. One "wild type" copy of the gene—that is, the most commonly occurring version—is enough to produce normal levels of the melanin polymer.